En mykorrhiza (fra gresk Myco , “sopp” og rhiza , “root”, et begrep som ble introdusert i 1885 av botanikeren Albert Bernhard Frank ) er resultatet av symbiotisk forening , kalt mycorrhization , mellom sopp og røtter av planter . Mycorrhizae er en hovedkomponent av edaphon og rhizosphere .
I denne generelt uspesifikke foreningen blir sporene til en mycorrhizal eller mycorrhizal sopp (fra gresk myco , rhiza og slekter , "til å fremkalle", bokstavelig talt som gir opphav til en mycorrhiza), spredt av vinden ( anemochoria ), av regn ( hydrohydrat ), eller av dyreskitt ( endozoochory ), spire, og gi opphav til hyfene i myceliet som koloniserer røttene til en plante. Disse rothyferne skiller seg fra ekstraradikulære hyfer som også vokser utenfor roten i flere centimeter, og utforsker jorden rundt vertsplantens rotsystem . Det som ofte kalles sopp, som er plukket med foten og hatten, er bare "fruktbarheten" av myceliet, sporoforen , der seksuell reproduksjon finner sted . Hyfer ser ut som fine filamenter, i stand til å utforske et veldig stort volum jord (tusen meter mycelfilamenter for en meter rot).
Mycorrhizal-forholdet er av den symbiotiske typen , men en ubalanse i forholdet kan induseres av en svakhet hos en av de to partnerne, foreningen kan da gli langs gjensidig-parasitismekontinuum. Soppen kan da også bidra til å resirkulere nekromassen til verten, til fordel for deres to etterkommere.
Mens leteoverflaten som brukes av planten, kun multipliseres med 10 av de aktive rothårene bare under spireperioden , multipliseres den med 10 000 takket være mycorrhizae (filamenter med en diameter på en hundredels millimeter) som har en energikostnad for å sette opp hundre ganger mindre enn røttene. Rotehårene forblir funksjonelle i noen få grupper av ikke-mykorrhizale voksne planter (mindre enn 10%) som sekundært har mistet soppforeningen, og mer enn 90% er kolonisert av disse hyfene av symbiontsoppen som gir mest mulig opptak av vann og næringsstoffer, og øke planteveksten ti ganger. Anslag antyder at det er nesten 50000 sopparter som danner mycorrhizal assosiasjoner med 250.000 plantearter, 80% av disse assosiasjonene er arbuskulær endomycorrhizae. Det ser ut til at ikke-mykorrhizale planter fra pionermiljøer (fravær av sopp), fuktige og / eller rike, der den hydrominerale tilførselen ikke krever mycorrhizae, "trengte for å kompensere for fraværet av sopphjelpen ved å utvikle rotstrukturer som etterligner mycelial filamenter og deres høye effektivitet i kolonisering av et stort volum jord ”.
Nåværende forskning viser en bredere forening på mycorrhizae-nivå, med begrepet mycorrhizal microbiome , kalt rhizomicrobiome, som også involverer bakterier.
Den opprinnelige symbionten ( det ser ut som en glomeromycete ) hadde dukket opp for rundt 500 millioner år siden i paleozoikumet , sannsynligvis samtidig med de første landplantene. De fossiler av floraen av Rhynie (f.eks aglaophyton (i) , Rhynia (i) ), i alderen ca. 400 millioner år inneholde morfologisk identisk til mycorrhizae Glomales , forbundet med jordstengler . Dette antyder at mycorrhizae var instrumentet for en akselerert kolonisering av fremkomne land, ved deres evne til å utvinne vann og mineraler fra jorden.
Disse første assosiasjonene var også i stand til å tillate dannelse av en jord som er bedre motstandsdyktig mot dårlig vær, bedre lagring av vann, samtidig som plantens motstand mot vannstress og kulde eller mangel på nitrogen (for eksempel i gran) forbedres , deres motstand mot vind og avrenning, og kanskje deres motstand mot overdreven lysintensitet, slik tilfellet er med mykofykobiontene til visse alger i fjæra (disse symbioser, med ascomycetes hovedsakelig, ser ut til å ha dukket opp sekundært).
Mycorrhizal assosiasjoner (arbuskulær og ektopomycorrhizal) eksisterer også i kalde økosystemer (gjennomsnittstemperatur under 15 ° C), der de er aktive hele eller deler av året (avhengig av jordtemperatur), involvert i visse tilpasninger av motstand mot gel og fremmer næringsinnsamling ved lav temperatur i mycorrhizal sopp. De bidrar utvilsomt til en bedre overlevelse av soppen i frossen jord om vinteren der mikrosoppene gjennomgår denne ekstra seleksjonsfaktoren. Disse kalde symbioses også fordelen gress som bygg ( Hordeum ), som dermed forbedrer dens adgang til jord fosfor.
Disse symbioene tilsvarer et ” makro- evolusjonært sprang ” (evolusjonært sprang) siden funksjonene til en partner er assosiert med funksjonene til den andre, med mangfoldige funksjonelle effekter. Faktisk passerer vi fra alge- og sopptilstanden til en jordbasert mykorrhizal plante (som strider mot darwinistisk gradualisme ).
For tiden er 85% av Archegoniates , så vel som Hepatic , endomycorrhizal av Glomales . Dette antar at symbiosen med Glomales er den eldste blant Archegoniates, og at den ville ha tillatt den imponerende strålingen til sistnevnte (mangfold, lignification ...). 80% av karplanter er kolonisert av endomycorrhizal sopp, mens ectomycorrhizae ofte er assosiert med treaktige trær.
De andre familiene til Glomeromycetes ( Acaulosporaceae og Gigaspora ) dukket opp senere for rundt -250 / -230 millioner år siden. De har overlegen kapasitet for utnyttelse av jordens mineralressurser.
De ectomycorrhizas , i mellomtiden, kan bli vist i kritt , men de eldste kjente fossiler dateres bare fra eocen . De tillot kolonisering av tidligere ugunstige jordarter. Det var også på dette tidspunktet at det kom nitrogenfikserende symbioser . Ectomycorrhizal symbiose har dukket opp flere ganger uavhengig av hverandre i forskjellige undertyper av sopp (80 ganger i Glomeromycetes , ascomyceter og basidiomyceter ) og Spermaphytes (12 ganger i Gymnosperms og angiospermer ). Denne frekvensen viser den evolusjonære suksessen til disse assosiasjonene fra forfedrene til ektomykorrhizalsopp som levde av dødt organisk materiale i jorden på en saprofytisk livsstil . Den komparative studien av genomene til nåværende ektomykorrhizal og saprofytisk sopp viser at mycorrhizalceller har mistet mange eksoenzymer som sørger for den autonome karbonholdige ernæringen til saprophytes, fordi de næres med sukker av vertsplanten, men har holdt enzymene (peroxidases, laccases, soppfenoloksidaser som tyrosinaser) som utnytter nitrogen- og fosfatressursene til organiske rester (lignin, fenolforbindelser, cellulose) som er nødvendige for at planten skal fungere korrekt.
Noen planter med nyere mykorrhizalsymbioser (fra et evolusjonært synspunkt) kan også inngå en tilknytning til arbuskulære mycorrhizae. Så det ser ut til at det har vært evolusjonære innovasjoner i denne typen symbiose. Disse innovasjonene forklarer utvilsomt det begrensede antallet planter som kan trekke dem sammen.
Utseendet til ectomycorrhizae har blitt korrelert to ganger med evolusjonær stråling fra planter:
Mer nylig Fortsatt andre former for endomycorrhizae og platoon ectomycorrhizae har dukket opp, spesielt i Ericales , med fenomenet mycoheterotrofi : deres soppsymbionter har enda sterkere saprofytisk kapasitet som tillater planten å innta organisk karbon igjen gjennom soppen, i jord der mineraliseringsaktiviteten er veldig lav ( myrer , høye fjell, kalde økosystemer, etc.): symbiosen tillater en kobling av trofiske nivåer .
Flere grupper av planter (mindre enn 10%) er ikke mycorrhizal. De har sekundært mistet assosiasjonen og illustrerer fenomenet neotenia . De er i hovedsak planter fra pionermiljøer (fravær av sopp) eller rike. For eksempel skum tilpasset fiendtlige miljøer, hvor de kan tørke ut og hvor en sopp ville overleve dårlig. De har skaffet seg anatomiske strukturer som tillater dem å motstå uttørking, men korrelativt er størrelsen fortsatt liten. Dette er også angiospermer ( Brassicaceae , Chenopodiaceae , Polygonaceae , Proteaceae , Caricaceae, etc.) som oftest har tilpasset seg rike miljøer, der den hydrominerale tilførselen ikke krever mycorrhizae.
Mycorrhizae er opprinnelsen til de mest komplekse økosystemene , og spesielt i skoger og spesielt tropiske skoger som ofte lever og utvikler seg på utakknemlige og noen ganger ufruktbare jordarter. Deres mycelium skjema koplet mycelie-nettverk som påvirker funksjonen av økosystemer (kretsløp, sammensetningen av plante fellesskap, karbonholdig næring av frøplanter i løpet av deres utvikling, modifisering av konkurranse, etc.) ved å tillate eller å øke betydelige strømmer av organisk karbon og mineraler ( nitrogen , fosfor , vann ...) via jorda (i gjennomsnitt 30 til 40% av mineralene fanget av margene i mycelialnettet, blir returnert til roten, sistnevnte gir 20 til 40% av karbohydratene som er fotosyntetisert til soppen). De utgjør et av de mest dynamiske elementene i mycorrhizal symbiose og spiller en viktig rolle i funksjon og strukturering av plantesamfunn. Disse overføringene er så effektive at de utfordrer konsentrasjonen av spesiering ved konkurranse om næringsstoffer mellom plantene i et økosystem, spesielt for fangst av fosfater ved røttene (de gjør det mulig å dispensere med fosfatgjødsel ), for motstand mot giftig kalsium (noen kalsikolplanter tåler kalsium takket være mycelial hyfer som aktivt avviser Ca 2+ -ioner utover eller ved å immobilisere dem i form av kalsiumoksalatkrystaller som faller ut i vakuoler , idioblaster eller på veggene til hyfer) eller for tørkebestandighet . De blir imidlertid fortsatt lite utnyttet i hagebruk , jordbruk og skogbruk , eller til dekontaminering av visse forurensede jordarter.
Enkelte soppgrupper er sannsynligvis nøkkelarter eller til og med “ingeniørarter” som påvirker de viktigste økologiske prosessene i jorden. De blir sett på av jordforskere som viktige elementer i mangfoldet i samfunn, noe som er en faktor for stabilitet og økologisk balanse. Mange nøkkelgrupper som finnes i jorden (spesielt bakterier og mycorrhizal sopp) kan koble seg til planter (minst 90% av terrestriske plantefamilier er bekymret) via arbuskulære mycorrhizalforeninger og spille synergier som er viktige for overlevelse og produktiviteten til planter et i hovedsak underjordisk økologisk nettverk , som noen biologer har kalt det trebrede nettet (med referanse til “ World wide web ”). I 1997 var den kanadiske forskeren Suzanne Simard den første som demonstrerte dette nettverket med mykorrhizal overføring av karbon mellom trær under naturlige forhold.
De fleste mycorrhizal sopper mistenkes for å ha flere verter eller til og med et bredt spekter av verter (hver plante er ofte assosiert med flere dusin forskjellige mycorrhizal sopp), som ser ut til å være bekreftet i naturlige miljøer , men studier gjort på dem Kultivert dyrkbar jord viser imidlertid , at mangfoldet av mykorrhizalsopp er "ekstremt lavt sammenlignet med skogjord" .
Koloniseringen av rotsystemer, det "mykorrhizogene" potensialet i jorden og "mycorrhizal avhengighet" av planter er omvendt korrelert med fosfationinnholdet i jordløsningen ; dessuten er dette resultatet ikke relatert til noen form for fosfatgjødsel , enten organisk eller uorganisk, siden planter bare absorberer ioner i oppløsning. Berikelsen av denne løsningen blir direkte ansvarlig for at den godt nærede planten ikke lenger fremmer utviklingen av mycorrhizae. I visse situasjoner blir oppnådde fosfornivåer uforenlige med installasjonen av mycorrhizae.
Mycorrhizae samhandler også med forskjellige jordbakterier (inkludert Pseudomonas ) som kan være patogene (mycorrhizae beskytter planten mot patogener, for eksempel ved å avgi antibiotika), men som også kalles "bakterier som er hjelpestoffer til mykorrhization" (på engelsk MHB: Mycorrhizal Helper Bacteria ) ettersom de spiller en så viktig rolle.
Mycorrhizae samhandler også med andre mycorrhizae og andre sopp og med visse luftrovdyr og parasitter av planter:
Mange kontrollerte mykorriseringseksperimenter har vist at naturlig eller kunstig regenerering har fordel av tilstedeværelse eller inokulering av symbiontsopp tilpasset plantene og konteksten. Omvendt kunne trøffelkultur bare eksporteres til New Zealand med mycorrhizal-vertsplanter. På samme måte vokste europeiske juletrær bare der med sine symbionter. Noen symbionter er spektakulære effektive: det er en økning på 60% av det totale volumet av Pseudotsuga menziesii som er tillatt på 10 år ved skogplanting fra ektopomykorrhizalplanter i planteskoler med en Laccaria bicolor- stamme kjent som S238N, sammenlignet med uinokulerte trær.
Forholdsregler : Det er en risiko for konkurranse med lokale arter, eller til og med genetisk forurensning . Flere oppfølginger har vist at planter som er inokulert i barnehager, generelt har mistet denne symbioten til andre fra lokal jord, men det kan ikke alltid være tilfelle.
I motsetning til hva mange tror, er mykorrhizal-forholdet ikke utelukkende symbiotisk fordi overføring av stoffer fra soppen til planten ikke alltid øker den selektive verdien (som er kriteriet for å definere symbiose ). Fordelene, som varierer veldig avhengig av genotypen til partnerne og miljøet, er vanskelige å vurdere, spesielt for flerårige planter. De kan dra nytte av symbiosen bare i korte perioder på forskjellige tidspunkter i livet. Det er faktisk et kontinuum av relasjoner mellom partnerne, fra symbiose til parasittisme gjennom saprotrofi (forholdet kan beskrives som en funksjon av kost / nytte-forholdet).
For eksempel er Tricholoma matsutake symbiotisk, parasittisk eller saprofytisk avhengig av utviklingsstadiet og miljøforholdene; orkide mycorrhizae er parasittisk på treaktige planter og fuglerede neotti er parasitic på mycorrhizae.
Faktisk er mykorrhisering kontinuerlig ustabil på grunn av interessekonflikter mellom partnere og seleksjonspresset som presser hver partner til å være så parasittisk som mulig, noe som forklarer de mange mulige interaksjonene: næringsutveksling (planten gir sukker og lipider , soppmycelia gir vann og mineralsalter som trekker fra bakken, noen ganger opptil flere meter fra roten), av vekstfaktorer ( hormoner , vitaminer ), og allomonene gir beskyttelse. For mye parasittisme kan føre til sammenbrudd. Over tid ville "symbiosen" ha dukket opp og ville ha blitt brutt flere ganger: assosiasjonen er reversibel. Saprofytiske sopp kan dermed være symbionter som har mistet verten. I visse mykorrhizale foreninger ser det ut til at en av partnerne utnytter den andre (enten av natur eller avhengig av forholdene). Uttrykket “utnytter” forstås her som et individ som oppnår en fordel (økning i selektiv verdi) uten gjensidighet. Vi kan nevne som et eksempel mycoheterotrofe planter (mer enn 200 arter kjent i orchidaceae , gentianaceae , burmanniaceae, etc.), planter med null eller redusert fotosyntetisk kapasitet som får karbon fra deres mycorrhizal-partner, og selv får det fra 'en klorofyllpartner gjennom mycelial-nettverket.
Utnyttelsen av en av partnerne til symbiosen av den andre antar kompensasjon for det utnyttede individet:
Kompensasjonsmekanismer kan være viktige for å stabilisere symbiosen. Generelt er symbioser preget av en reduksjon i genetisk drift og spekteringshastigheten (Jf. "Red King" -effekten ), men dette er ikke alltid tilfelle ved mykorrhisering: jo mer assosiasjonen er utnyttende, jo mer er den spesifikk fordi den utnyttet utvikler motstand som må omgåes av utbytteren ( The Red Queen ). I det foreliggende tilfelle ville det selektive trykket på verten ført til lignin og favorisert veksten av vaskulært vev.
Til slutt antyder logikk og mange ledetråder en parasittisk opprinnelse til mycorrhizae:
Det er to hovedtyper av mycorrhizae, definert av de fysioanatomiske forholdene mellom de to partnerne: endomycorrhizae og ectomycorrhizae. Ericoid mycorrhizae og de som er forbundet med orkideer blir også studert for deres økologiske tjenester , men har en mer begrenset økonomisk interesse.
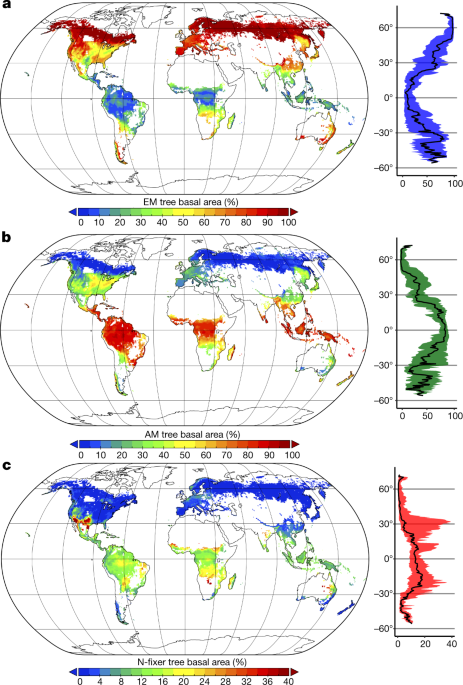
En global kartlegging av skogssymbioser viser ektomykorrhizaltræres evolusjonære suksess . Av de tre mest vanlige typene symbiose (arbuscular endomycorrhizae rikelig i varmt og fuktig tropiske klimaer, ectomycorrhizae i kaldt klima, og nitrogen- feste bakterier i tørre og varme klima), representerer ectomycorrhizae bare 2% av plantearter, men utgjør 60% av alle trær på planeten.
Endomycorrhizae (eller indre mycorrhizae) er den vanligste formen. Dette er mycorrhizae som trenger inn i røttene for å bedre assosiere seg med dem.
Det er flere typer endomycorrhizae:
De er assosiert med urteaktige og treaktige planter. Disse arbuskulære endomycorrhizae, også kalt vesikulær og arbuskulær mycorrhizae (eller vesiculo-arbuskulær mycorrhizae), får navnet sitt fra intercellulære vesikler (ofte dråper olje, reservelagring for soppen) og "intracellular" strukturer som minner om et lite tre. Hvis de krysser veggen godt, trenger de imidlertid ikke inn i plasmamembranen i plantecellen, innhold som forårsaker invaginasjon av membranen derav. Dette har den effekten at kontaktflaten mellom hyfa og plantecellen økes og dermed letter utveksling av metabolitter mellom de to partnerne.
Arbuscular endomycorrhizae dannes bare av sopp fra divisjonen Glomeromycetes som har mistet seksuell reproduksjon. Hyfene strekker seg inn i kortikalt parenkym av roten, og danner vesikler som inneholder reserver, og forgreningsstrukturer, arbuskulene. De reproduserer derfor bare aseksuelt. Imidlertid kan hyfer fra forskjellige individer smelte sammen, noe som muliggjør genetisk utveksling og en form for paraseksualitet.
De er også unike genetisk siden sporene har flere genetisk forskjellige kjerner.
I denne symbiosen hjelper det tette og omfattende nettverket av hyfer av mykorrhizalsopper planten å utforske et økt volum jord og få tilgang til steder som er utilgjengelige for røttene. Soppen lar planten forbedre ernæringen ved å tilveiebringe hovedsakelig vann, fosfor og nitrogen . For øvrig gjør koloniseringen av røttene av mycorrhizal sopp det mulig å beskytte dem mot angrep fra patogene organismer. Flere studier har vist at uten tilknytning til en symbiotisk sopp, vil planten vokse saktere (hvis i det hele tatt, som i tilfelle svart furu på kalkholdig jord) og vil være mye mer sannsynlig å bli offer for en infeksjon. Til gjengjeld drar soppen fordel av fotosyntese av planten i form av organisk materiale som er rik på energi ( sukker ) som er viktig for dens overlevelse. Kvaliteten på humus forbedres derved, til fordel for andre arter og for vedlikehold eller konstitusjon og forbedring av jorden.
Noen planter av Gymnosperm- familien, som Podocarpaceae , Araucariaceae og Phyllocladaceae , er kjent for å ha rothevelser invadert av endomycorrhizal sopp, kalt myconodules eller pseudonodules .
De ectomycorrhizas (eller mycorrhizae ekstern) omfatter kun 5% av karplanter, for det meste trær av temperert og barskogs (for eksempel Fagaceae , den Pinacées eller Betulaceae ) og sopp av delingen av Ascomycetes , de Basidiomycetes eller Zygomycetes . Disse mykorrhizene trenger ikke gjennom cellevegger inne i plantens celler, men bare omgir røttene og danner en kappe av mycelium og et nettverk mellom celleveggene til roten.
Soppen knytter seg først til fine røtter med bestemt vekst, uten rothår. Deretter omslutter den roten til en kappe av hyfer , mycorrhizal-ermet. Andre hyfer vokser mellom celler i den ytre delen av kortikalt parenkym og danner det symbiotiske grensesnittet eller " Hartig-nettverket ". Symbiose endrer fysiognomien til mycorrhizal-roten: den svulmer, slutter å vokse og kan forgrene seg rikelig. Hetten og den apikale meristen reduseres.
Hundrevis av forskjellige sopper kan knyttes til samme art. Den bøk , for eksempel, har en "record" med mer enn 200 mychoriziennes foreninger.
Veksten av soppmyceliet mot røttene til vertsplanten utløses av oppfatningen av et signal som kontinuerlig sendes ut av røttene i form av molekyler som er spesifikke for planten, slik som strigolaktoner , og spesielt flavonoider .
Signalene som sendes ut av soppen som lar den bli gjenkjent av planten, er dårlig forstått. I analogi med Nod-faktorene kalles disse faktorene "Myc-faktorer". Nodfaktormolekyler produsert av arbuskulære sopp og som har en effekt på vertsplanten har nylig blitt demonstrert, men deres rolle i symbiotisk signalering gjenstår å bli bestemt.
Den myc-faktoraktiverte signalveien har trinn til felles med Nod-banen som er tilstede i belgfrukter som er nodulert av rhizobia og i aktinorrhizale planter . Reseptorene til Myc-faktorene er ikke kjent, men proteinene som griper inn etterpå som SYMRK, CASTOR, POLLUX, NUP, CYCLOPS er nødvendige for de rhizobiale og arbuskulære mykorrhizale symbioser. Som med rhizobial symbiose induserer kontakt med en arbuskulær sopp kalsiumsvingninger. Da arbuskulær endomykorrhizae dukket opp før nitrogenfikserende endosymbiose, antar forskere at mykorrhizaltransduksjonssveien ble rekruttert av nitrogenfikserende bakterier.
En optimal mycorrhizal-prosesjon vil fungere som en "gjødsel" uten ulempene, og uten kostnad, uten plantevernmidler , soppdrepende midler og endringer . Etter innledende forsøk i Australia på 1920-tallet utviklet selskaper inokuleringsstammer som var egnet for flere kommersielle arter (enkelt- eller multistamme-inokula), men de måtte også tilpasses konteksten til stedet. Flere forskere mener at stammene av genetisk symbiontsopp som er best egnet til åkeren, er de som er urfolk. Private selskaper over hele verden har således utviklet produksjon og salg av mykorrhizale inokulanter beregnet til landbruksbruk ( biogjødsel , biostimulerende middel ), men også økologiske ( revegetasjon av erodert jord, fytoremediering ).
I landbruket ville bruk av mykorrhizal sopp unngå mange problemer med denaturering av organisk jord. Tilsetning av plantevernmidler og spesielt soppdrepende midler kan faktisk ha skadelige effekter på jordkvaliteten. Sopp har viktige roller i å opprettholde fruktbar jord. Tilsetningen av sterkt fosforholdige kjemiske forbindelser ville indusere en utarmning av mengde og kvalitet av endomykorrhizalsopp, og redusert tilførsel av næringsstoffer til dyrkede planter. I tillegg har for mye fosfor en negativ innvirkning på vannveier (eutrofiering). Det er derfor viktig å redusere mengden av skadelige kjemikalier og i stedet velge inokulering av naturlige sopper som vil ha de samme fordelene med rask vekst, uten ulempene for miljøet.
En studie på jujubetre viste at kontrollert mykorrhisering var gunstig for vekst og fosfat ernæring. Faktisk absorberte frukttreet lettere fosfor tilsatt i dårlig jord. Han var til og med avhengig av det for normal vekst. Det er derfor ikke nok i landbruket å bare tilsette fosfor i veldig store mengder for at planten skal vokse raskere, det er også et spørsmål om å ta hensyn til mycorrhizal-nettverket. Den må først og fremst kunne absorbere effektivt. En overdose av næringsstoffer vil ikke endre noe, og dette vil bare føre til utvasking av fosfor og forurensning av overflatevann.
Handlingen av mycorrhizae stopper ikke med bedre ernæring: de lar også planten bedre beskytte seg mot skadelige biotiske og abiotiske faktorer. Selvfølgelig innebærer bedre ernæring bedre plantehelse, som da bedre tåler miljøbelastninger, spesielt patogener. Mycorrhizae kan frigjøre antibiotika i rhizosfæren som har den effekten at de eliminerer alle mikroorganismer, patogene eller ikke, noe som kan avlede deler av handelen til deres fordel. I tillegg induserer mykorrhizalsymbiose etablering av forsvarsmekanismer i selve planten (immunitet og immunstimulering). Det er en indirekte beskyttelse som ligger på mobilnivå. I ectomycorrhizal planter , rhizoderm cellene syntetisere tanniner som er lagret i vakuoler som gir beskyttelse mot ytterligere angrep av mikrober og andre patogener. Anlegget produserer også mer lignin i celleveggene i endoderm og vaskulære vev. Det kan også indusere eller undertrykke forskjellige forsvar knyttet til fytoalexiner , peroksidaser , kitinaser og flere andre. Kort sagt forårsaker soppen betydelige endringer i forsvarsmekanismene i planten mot forskjellige parasitter, og dette før infeksjoner.
Noen ganger forårsaker tilknytningen til en sopp en ombygging av rotsystemet. For eksempel forårsaker mykorrhisering hos jordbær økt beskyttelse mot rotrot forårsaket av Phytophthora fragariae . Den intense forgreningen av røttene som er fremkalt av mycorrhizal sopp er ledsaget av roteksudater, og dette vil ha effekten av å endre mikrofloraen og de direkte interaksjonene med parasittene.
Når det gjelder parasittiske sopp, er en plante assosiert med en mycorrhiza også bedre beskyttet. Hyfene til den symbiotiske soppen koloniserer plantens røtter og "blokkerer" tilgangssteder for den parasittiske soppen. De konkurrerer også med næringsstoffene som er tilgjengelige. En mycorrhiza får også fordeler ved å binde seg til en plante: den mottar karbonforbindelser i bytte mot mineraler. Siden karbon er svært ettertraktet, har en mycorrhizae en større fordel enn en parasittisk sopp fri i jorda. I tillegg kan mycorrhizae utgjøre opptil 80% av jordens mikrobielle masse. De påvirker derfor de fysiske og kjemiske egenskapene til miljøet sterkt og kan derfor kontrollere flere mikrobielle interaksjoner i jorden. Noen ganger er det i samsvar med mykorrhizalplanten ved å gi beskyttelse og vekst, men noen ganger er effektene ganske negative.
Mycorrhizae kan derfor bidra til å beskytte planten mot abiotiske (f.eks. Tørke) og biotiske belastninger på flere måter: bedre ernæring og helse, morfologisk transformasjon av røttene, induksjon eller undertrykkelse av forsvarsmekanismer og ved å handle på selve parasitten, enten ved konkurranse om ressurser og infeksjonssteder, eller ved modifisering av mikrofloraen og en økning i nivået av organisk materiale.
Det skal imidlertid bemerkes at disse interaksjonene studeres i drivhus og i kontrollerte miljøer, og at de derfor ikke fullt ut reflekterer all kompleksiteten i et naturlig miljø som varierer i tid og i sammensetning.
Med alle de oppgitte fordelene med mycorrhizae, er det derfor rimelig å si at riktig bruk av dem ville unngå mange miljøhensyn, da de ville fungere som gjødsel og beskyttelsesmidler. Bruk av kjemisk gjødsel, soppdrepende midler og plantevernmidler ville da ikke være like nødvendig. Gjødsel reduserer faktisk mykorriseringshastigheten, noe som har effekten av økt avhengighet av planter av disse inngangene. Likeledes reduserer bruken av plantevernmidler jordens inokuleringskraft, noe som resulterer i en reduksjon i den beskyttende effekten av mycorrhizae og et økt behov fra planter for plantevernmidler. Tillegget av slike innganger setter opp en form for ond sirkel .
Skogbruk og landbruksaktivitet kan forstyrre eller modifisere (negativt eller positivt) soppmikrofloraen og dens evne til å mykorrisere;
Biokjemiske og biologiske endringer, så vel som mikroklimatiske forandringer som følge av store klare kutt, (eller til og med høst i landbruket) kan ha undervurdert påvirkninger, knyttet til direkte påvirkning på jorden, på vann, men også til tap av sopp inokulum (selv om røttene forblir i jorda, blir " stort tre " og stort " dødt tre " sjeldne eller fraværende, og skråstrek blir ofte samlet).
Begrepet mycorrhizae (fra gresk myco , "sopp" og rhiza , "rot") ble introdusert i 1885 av botanikeren Albert Bernhard Frank . Ansatt av den prøyssiske kongen William jeg st å utvikle metoder for å produsere trøfler ved å studere hvordan de sprer seg, konstaterer ABFrank at disse sopp forbinder med røtter av trær og foreslår at denne foreningen er et symbiotisk forhold og ikke parasittisk . Franks teori om evolusjonær økologi , som motsier troen på at trøfler og andre sopper forårsaker plantesykdom og råtn, bestrides av kollegene. Denne teorien vender tilbake til sentrum med arbeidet til Jack Harley som betraktet som "paven til mycorrhizae" for studiene siden 1950-tallet, Barbara Mosse i 1962, JM Phillips og DS Hayman i 1970, M. Gardes og TD Bruns i 1993.
Forskning er i rask utvikling i begynnelsen av XXI th århundre med nye former for molekylærbiologi og genetikk til poenget med å gi navn til en vitenskapelig disiplin , det mycorhizologie , studert av mycorhizologues . Fra 2008 til 2016 gjaldt mer enn 10.000 nye vitenskapelige bidrag mycorrhizae.
{kind=link}